Ревизия рода Ariocarpus
Edward F. Anderson, W. A. Fitz Maurice
Haseltonia 5:1-20.(1997)
 Резюме. Авторы предлагают рассматривать
A. fissuratus var. hintonii, как подвид
A. bravoanus. Они предлагают упразднить подроды
Ariocarpus — подрод Ariocarpus и подрод
Roseocactus (Aegopadothele), также же, как и внутривидовые
таксоны A. fissuratus var. fissuratus и
var. lloydii. Они соглашаются, что Ariocarpus
(Anhalonium) elongatus является синонимом A.
retusus. Основываясь на факте широкого совместного разделения
характерных особенностей и недавней идентификации в большой гибридной массе
между A. retusus и A. trigonus, авторы
заключают, что A. trigonus — подвид A.
retusus. Новый ключ видов и разновидностей отражающий предложения
приведенные выше — включен. Статус сохранности видов в
пределах рода Ariocarpus попеременно классифицируется от «В
опасности» до «Низкий риск».
Резюме. Авторы предлагают рассматривать
A. fissuratus var. hintonii, как подвид
A. bravoanus. Они предлагают упразднить подроды
Ariocarpus — подрод Ariocarpus и подрод
Roseocactus (Aegopadothele), также же, как и внутривидовые
таксоны A. fissuratus var. fissuratus и
var. lloydii. Они соглашаются, что Ariocarpus
(Anhalonium) elongatus является синонимом A.
retusus. Основываясь на факте широкого совместного разделения
характерных особенностей и недавней идентификации в большой гибридной массе
между A. retusus и A. trigonus, авторы
заключают, что A. trigonus — подвид A.
retusus. Новый ключ видов и разновидностей отражающий предложения
приведенные выше — включен. Статус сохранности видов в
пределах рода Ariocarpus попеременно классифицируется от «В
опасности» до «Низкий риск».
Введение
Более чем 35 лет назад один из нас (Андерсон) опубликовал серию
статей, посвященных таксономии и экологии рода Ariocarpus
(Андерсон, 1958, 1960, 1962, 1963, 1964). Со времени тех работ, было много
написано относительно этого рода и было открыто несколько новых таксонов.
Мы также смогли изучить много дополнительных популяций Ariocarpus
в поле, включая все те таксоны, которые были предложены в течении последних
35 лет. Эта статья представляет новые данные и толкования рода,
основанные на наших недавних лабораторных работах, изучения растений в поле,
и анализа литературы.
наверх
Таксономическое состояние A. bravoanus и A. fissuratus var. hintonii
 Два новых таксона Ariocarpus были описаны в пределах
прошлых семи лет. Ступпи и Тейлор (1989) описали новый таксон Ariocarpus.
Оригинальное «открытие» этого растения произошло в экспериментальном
заказнике около Матехуала, S.L.P., работающего под руководством Universidad
Autonoma Agzipia — Антонио Нарро, Салтилло, Коахуила.
Смотритель продавал собранные растения, которые включали новый таксон.
Несколькими годами позже, Джордж Хинтон обнаружил место произрастания этого
растения приблизительно на 25 километров южнее экспериментального сада. Один
из нас (Фитц Морис), вместе с Томом Дэвисом, искал дополнительные популяции
растений и нашел такое место около экспериментального заказника (Фитц Морис
и Дэвис, 1987). В то время, местные жители рассказали о существенном сборе
растений на том самом месте иностранцами. Это почти то же самое местообитание,
которое было ранее зафиксировано смотрителем в этом экспериментальном заказнике.
Этот новый таксон, казалось, наиболее близко походил на A.
fissuratus — широко распространенный и высоко
изменчивый вид, растущий во многих местах от северной Мексики и до Техаса.
Растения, были отданы одним из нас (Фитц Морис), Ступпи и Тейлору (1989),
которые описали его как A. fissuratus var.
hintonii.
Два новых таксона Ariocarpus были описаны в пределах
прошлых семи лет. Ступпи и Тейлор (1989) описали новый таксон Ariocarpus.
Оригинальное «открытие» этого растения произошло в экспериментальном
заказнике около Матехуала, S.L.P., работающего под руководством Universidad
Autonoma Agzipia — Антонио Нарро, Салтилло, Коахуила.
Смотритель продавал собранные растения, которые включали новый таксон.
Несколькими годами позже, Джордж Хинтон обнаружил место произрастания этого
растения приблизительно на 25 километров южнее экспериментального сада. Один
из нас (Фитц Морис), вместе с Томом Дэвисом, искал дополнительные популяции
растений и нашел такое место около экспериментального заказника (Фитц Морис
и Дэвис, 1987). В то время, местные жители рассказали о существенном сборе
растений на том самом месте иностранцами. Это почти то же самое местообитание,
которое было ранее зафиксировано смотрителем в этом экспериментальном заказнике.
Этот новый таксон, казалось, наиболее близко походил на A.
fissuratus — широко распространенный и высоко
изменчивый вид, растущий во многих местах от северной Мексики и до Техаса.
Растения, были отданы одним из нас (Фитц Морис), Ступпи и Тейлору (1989),
которые описали его как A. fissuratus var.
hintonii.
 Только несколькими годами позже после публикации A.
fissuratus var. hintonii, Гектор Эрнандез обнаружил
новую и отличающуюся популяцию Ariocarpus, растущую намного южнее
в штате Сан Луис Потоси
(Карта 1).
Это растение было описано Эрнандезом и
Андерсоном (1992) как A. bravoanus.
Только несколькими годами позже после публикации A.
fissuratus var. hintonii, Гектор Эрнандез обнаружил
новую и отличающуюся популяцию Ariocarpus, растущую намного южнее
в штате Сан Луис Потоси
(Карта 1).
Это растение было описано Эрнандезом и
Андерсоном (1992) как A. bravoanus.
 Летом 1993 мы посетили местобитание этих двух новых таксонов в
компании Джорджа С. Хинтона. Ariocarpus fissuratus var.
hintonii теперь известен только из двух местностей на высоте
приблизительно 1400 метров, разделенных небольшим пространством
приблизительно в 25 километров. Они растут в местности типичной для пустыни
Чихуахуа, в штате Сан Луис Потоси
(Карта 1).
Множество растений были найдены в обоих местах, хотя они были хорошо
замаскированы в почве.
Ariocarpus bravoanus известен только из единственного места
произрастания на высоте 1500 метров в несколько измененной пустыне
Чихуахуа, также в пределах штата Сан Луис Потоси.
Летом 1993 мы посетили местобитание этих двух новых таксонов в
компании Джорджа С. Хинтона. Ariocarpus fissuratus var.
hintonii теперь известен только из двух местностей на высоте
приблизительно 1400 метров, разделенных небольшим пространством
приблизительно в 25 километров. Они растут в местности типичной для пустыни
Чихуахуа, в штате Сан Луис Потоси
(Карта 1).
Множество растений были найдены в обоих местах, хотя они были хорошо
замаскированы в почве.
Ariocarpus bravoanus известен только из единственного места
произрастания на высоте 1500 метров в несколько измененной пустыне
Чихуахуа, также в пределах штата Сан Луис Потоси.
 Эта популяция состоит не больше, чем из нескольких тысяч растений, ограниченных известной
областью меньшей одного квадратного километра. Единственными другими
видами Ariocarpus, встречающимися в этой области являются —
A. retusus (в сходных местах произрастания) и A.
kotschoubeyanus (растущий в очень отличающихся местах, обычно на
наносных (илистых) плато). Ariocarpus аgavoides, который
Эрнандез и Андерсон (1992) предложили считать близко связанным с A.
bravoanus, встречается на расстоянии более чем в 130 километров далее
в штате Тамаулипас.
(Карта 2).
Ariocarpus fissuratus var. hintonii происходит
из штата Сан Луис Потоси на расстоянии 85 километров.
К сожалению, живыми растениями Ariocarpus fissuratus
var. hintonii ни Эрнандез, ни Андерсон не располагали во время
их описания A. bravoanus, и при этом они не видели их в
природе.
Эта популяция состоит не больше, чем из нескольких тысяч растений, ограниченных известной
областью меньшей одного квадратного километра. Единственными другими
видами Ariocarpus, встречающимися в этой области являются —
A. retusus (в сходных местах произрастания) и A.
kotschoubeyanus (растущий в очень отличающихся местах, обычно на
наносных (илистых) плато). Ariocarpus аgavoides, который
Эрнандез и Андерсон (1992) предложили считать близко связанным с A.
bravoanus, встречается на расстоянии более чем в 130 километров далее
в штате Тамаулипас.
(Карта 2).
Ariocarpus fissuratus var. hintonii происходит
из штата Сан Луис Потоси на расстоянии 85 километров.
К сожалению, живыми растениями Ariocarpus fissuratus
var. hintonii ни Эрнандез, ни Андерсон не располагали во время
их описания A. bravoanus, и при этом они не видели их в
природе.
 Лишь недавно мы получили возможность наблюдать естественные вариации
в популяциях A. bravoanus и Ariocarpus fissuratus
var. hintonii, также как препарировать несколько экземпляров,
чтобы изучить строение сосков и ареол и исследовать семена с растровым электронным
микроскопом, чтобы вывести некоторые заключения относительно вероятных соотношений
этих растений к другим таксонам Ариокарпусов. Эрнандез и Андерсон (1992) высказывались
о том, что A. bravoanus «обладает комбинацией морфологических
признаков, не соответствующих любой из известных разновидностей этого рода»
и что его «признаки, фактически, размещают его как промежуточное звено
между родами Ariocarpus и Roseocactus». Наши исследования
1993-96, укрепляют это утверждение и ведут нас к иному толкованию
идеям Андерсона (1960, 1962) относительно развития ареол у Ариокарпусов.
Лишь недавно мы получили возможность наблюдать естественные вариации
в популяциях A. bravoanus и Ariocarpus fissuratus
var. hintonii, также как препарировать несколько экземпляров,
чтобы изучить строение сосков и ареол и исследовать семена с растровым электронным
микроскопом, чтобы вывести некоторые заключения относительно вероятных соотношений
этих растений к другим таксонам Ариокарпусов. Эрнандез и Андерсон (1992) высказывались
о том, что A. bravoanus «обладает комбинацией морфологических
признаков, не соответствующих любой из известных разновидностей этого рода»
и что его «признаки, фактически, размещают его как промежуточное звено
между родами Ariocarpus и Roseocactus». Наши исследования
1993-96, укрепляют это утверждение и ведут нас к иному толкованию
идеям Андерсона (1960, 1962) относительно развития ареол у Ариокарпусов.
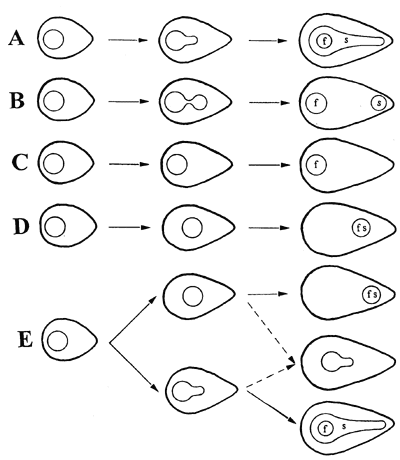
Рис. 5
Как показано на рисунке 5, Андерсон (1960, 1962) описал четыре схемы
развития ареол у Ариокарпусов.
A) A. fissuratus и A. kotschoubeyanus,
у которых удлинение происходит ниже ареолы, таким образом, создавая удлиненную
структуру внутри бороздки;
B) A. retusus, у которого несущие колючки и цветки части
ареолы разделяются в процессе удлинения, создавая диморфные ареолы;
C) A. trigonus и A. scaphirostris, у
которых удлинение сосочка происходит вне ареольной области;
D) A. agavoides, у которого удлиняется часть сосочка,
расположенная в основании неделимой ареольной области, но после того, как
цветение произошло на молодом сосочке, подобно всем видам, в центре растения.
Андерсон (1960) отметил, что иногда наблюдаются отклонения от этих
схем, продемонстрировав в качестве примера экземпляры A. fissuratus,
у которых отсутствуют ареолы в виде бороздки. Тем не менее, он считал,
что между видами имеющими ареолы в форме бороздки и теми, у которых их нет,
имелись все же достаточные различия для того, что бы разделить их на подрода.
Мы теперь поставим под вопрос это решение и обсудим его в следующем разделе
этой статьи.
 Новые свидетельства говорят, что схемы B и C не всегда различны и
что иногда индивидуальные растения могут иметь оба этих типа; это обсуждено
далее в дальнейших разделах. Также, к четырем схемам, описанным выше, должна
быть добавлена пятая, представляющая ареолы развивающиеся у двух недавно описанных
таксонов — A. bravoanus и A.
fissuratus var. hintonii. Однако эта схема
значительно более усложнена, чем наблюдаемые в других групах. Как показано в
Схеме E на Рисунке 5, сосочек может развиваться (и удиняться) либо позади либо
ниже ареольной области, развиваясь в состояние подобное A или D. И снова,
удлинение происходит прежде всего после цветения. Промежуточные состояния также
встречаются, создавая ареольные области в середине соска, с различными степенями
удлинения.
Новые свидетельства говорят, что схемы B и C не всегда различны и
что иногда индивидуальные растения могут иметь оба этих типа; это обсуждено
далее в дальнейших разделах. Также, к четырем схемам, описанным выше, должна
быть добавлена пятая, представляющая ареолы развивающиеся у двух недавно описанных
таксонов — A. bravoanus и A.
fissuratus var. hintonii. Однако эта схема
значительно более усложнена, чем наблюдаемые в других групах. Как показано в
Схеме E на Рисунке 5, сосочек может развиваться (и удиняться) либо позади либо
ниже ареольной области, развиваясь в состояние подобное A или D. И снова,
удлинение происходит прежде всего после цветения. Промежуточные состояния также
встречаются, создавая ареольные области в середине соска, с различными степенями
удлинения.
Дальнейшие исследования двух новых таксонов приводят к предположению,
что они более близко связаны друг с другом, чем с другими любыми таксонами
Ариокарпусов. Не только их область распространения близка друг к другу, чем к
другим возможным близким формам, типа A. fissuratus или A.
agavoides, но они разделяют набор сходных характеристик. Они миниатюрны
в размере, вероятно признак неотении, подобное также замечено у A.
agavoides и A. scaphirostris. Они оба обладают
эпидермальными бугорками в различной степени (Рисунки 7-10); которые отсутствуют
у всех других таксонов кроме A. fissuratus. Сканирующий
электронный микроскоп показывает, что их семена схожи и по размерам и по
кутикулярной природе бородавчатой тесты (Эрнандез и Андерсон, 1992), но они
определенно отличаются от семян A. fissuratus. За исключением
A. retusus и A. kotschoubeyanus, которые
встречаются во множестве мест на севере и юге, A. bravoanus
и A. fissuratus var. hintonii являются
самыми южными таксонами рода. Эти таксоны встречаются в пределах примерно 85
километров (50 миль) друг от друга, но они удаленны от ближайшего
известного местоположения A. fissuratus —
более чем на 200 километров (120 миль).
 Они также имеют схожие
сосочки и строение цветка по сравнению с другими таксонами Ариокарпусов.
Они разделяют тенденцию вертикальной ориентации и размера сосочка, их мелкие
бугорки (папиллы) также имеют общие сходные характеристики для этих двух
таксонов и отличающиеся от A. fissuratus. И, наконец,
исследование индивидуальных экземпляров растений всех известных популяций
показывает промежуточные формы, особенно в отношении развития ареол. Некоторые
экземпляры A. bravoanus, например, имеют частично развитую
бороздку на ареолах, тогда как у некоторых экземпляров
A. fissuratus var. hintonii эта бороздка
почти отсутствует.
Они также имеют схожие
сосочки и строение цветка по сравнению с другими таксонами Ариокарпусов.
Они разделяют тенденцию вертикальной ориентации и размера сосочка, их мелкие
бугорки (папиллы) также имеют общие сходные характеристики для этих двух
таксонов и отличающиеся от A. fissuratus. И, наконец,
исследование индивидуальных экземпляров растений всех известных популяций
показывает промежуточные формы, особенно в отношении развития ареол. Некоторые
экземпляры A. bravoanus, например, имеют частично развитую
бороздку на ареолах, тогда как у некоторых экземпляров
A. fissuratus var. hintonii эта бороздка
почти отсутствует.
 Мы заключаем, что два новых таксона представляют единственный вид,
с промежуточными чертами между A. agavoides и A.
fissuratus. Мы предлагаем, что бы внутривидовой таксон hintonii
был сохранен, для известных популяций этого таксона географически изолированных
от любых других популяций Ариокарпусов. Если в дальнейших полевых работах смогут
найтись новые популяции этого вида размещенные в промежутке между ныне известным,
и с экземплярами демонстрирующими даже большую гетерогенность сосочков и характера
ареол то, возможно, что внутривидовой таксон не должен быть формально признанным.
Это сделало бы A. bravoanus схожим с A. fissuratus
и A. retusus, которые являются широко распространенными видами
с большими вариациями в строении сосочков (смотри следующие разделы). При этом,
мы предлагаем следующую новую комбинацию, веря, что она лучше всего демонстрирует
наше нынешнее понимание взаимоотношений внутри этого рода: Ariocarpus
bravoanus H. Hernandez & E.F. Anderson subsp. hintonii
(W. Stuppy & N.P. Taylor) F.F. Anderson & W.A. Fitz Maurice, comb. et stat nov.
(Basionym = Ariocarpus fissuratus var. hintonii W. Stuppy & N.P. Taylor,
Bradleya 7:84. 1989).
Мы заключаем, что два новых таксона представляют единственный вид,
с промежуточными чертами между A. agavoides и A.
fissuratus. Мы предлагаем, что бы внутривидовой таксон hintonii
был сохранен, для известных популяций этого таксона географически изолированных
от любых других популяций Ариокарпусов. Если в дальнейших полевых работах смогут
найтись новые популяции этого вида размещенные в промежутке между ныне известным,
и с экземплярами демонстрирующими даже большую гетерогенность сосочков и характера
ареол то, возможно, что внутривидовой таксон не должен быть формально признанным.
Это сделало бы A. bravoanus схожим с A. fissuratus
и A. retusus, которые являются широко распространенными видами
с большими вариациями в строении сосочков (смотри следующие разделы). При этом,
мы предлагаем следующую новую комбинацию, веря, что она лучше всего демонстрирует
наше нынешнее понимание взаимоотношений внутри этого рода: Ariocarpus
bravoanus H. Hernandez & E.F. Anderson subsp. hintonii
(W. Stuppy & N.P. Taylor) F.F. Anderson & W.A. Fitz Maurice, comb. et stat nov.
(Basionym = Ariocarpus fissuratus var. hintonii W. Stuppy & N.P. Taylor,
Bradleya 7:84. 1989).
наверх
Состояние подрода Ariocarpus
Бергер (1925) обосновал род Roseocactus как отдельный от
рода Ariocarpus, аргументируя это тем, что их ареолы имеют совершенно
различные типы строения и что место возникновения цветка у них различно.
Андерсон (1960) принял первый аргумент Бергера, но продемонстрировал, что
фактически, место возникновения цветка было, по существу, тем же самым у всех
видов Ariocarpus (включая Roseocactus). Поэтому он принял
классификацию В. T. Маршалла (1946) и понизил ранг Roseocactus до
подрода. Хант и Тейлор, правильно указали в редакционном примечании к работе
Эрнандеза и Андерсона (1992), что правильное название для подрода Roseocactus
(A. Berger) W.T. Marshall должно быть — подрод Aegopodothele
K. Schumann, поскольку это было опубликовано им 48-ю годами
ранее (Шуман, 1898).
В дополнительном исследовании правильности разделения между двумя
подродами Ariocarpus, как бы они не были бы названы, мы заключаем:
что не имеется больше никакого оправдания для такого раздвоения, принимая во
внимание строение ареол у A. bravoanus. Очевидно, что не
имеется никаких оснований для мнений о том, что имеются два отличающихся типа
строения ареол в роде, потому что A. bravoanus и его подвид
hintonii имеют многочисленные промежуточные формы.
наверх
Внутривидовые таксоны A. fissuratus
 Андерсон (1963) и другие (Бартлотт, 1979; Хант, 1989; Х. Браво и
Санчез-Мехорада, 1991) признавали две разновидности A. fissuratus
var. fissuratus и var. lloydii.
Ступпи и Тейлор (1989) предложили var. hintonii
как третью разновидность A. fissuratus, но в предыдущем
разделе этой статьи мы рекомбинировали его как подвид A. bravoanus.
Андерсон (1963) и другие (Бартлотт, 1979; Хант, 1989; Х. Браво и
Санчез-Мехорада, 1991) признавали две разновидности A. fissuratus
var. fissuratus и var. lloydii.
Ступпи и Тейлор (1989) предложили var. hintonii
как третью разновидность A. fissuratus, но в предыдущем
разделе этой статьи мы рекомбинировали его как подвид A. bravoanus.
 Можно легко увидеть различия между растениями из большинства северных
популяций в округе Биг Бенд (Техас) — var.
fissuratus — и таковых же из южных популяций
— var. lloydii — растущих около
Паррас (Коахуила) и встречающихся западнее Назас (Дюранго). Ступпи и Тейлор
(1989) в описании новой разновидности hintonii обсуждали характеристики
и распространение двух разновидностей A. fissuratus так же,
как и проблемы промежуточных форм. Они пришли к заключению, что «эти две
разновидности могут отличаться намного более чем их ареалами произрастания и
кажутся стоящими сохранения».
Можно легко увидеть различия между растениями из большинства северных
популяций в округе Биг Бенд (Техас) — var.
fissuratus — и таковых же из южных популяций
— var. lloydii — растущих около
Паррас (Коахуила) и встречающихся западнее Назас (Дюранго). Ступпи и Тейлор
(1989) в описании новой разновидности hintonii обсуждали характеристики
и распространение двух разновидностей A. fissuratus так же,
как и проблемы промежуточных форм. Они пришли к заключению, что «эти две
разновидности могут отличаться намного более чем их ареалами произрастания и
кажутся стоящими сохранения».
 Мы, также, были обеспокоены присутствием промежуточных форм
A. fissuratus от Куатро Синегас до Эстасион Марте в штате
Коауила. Кроме того, многие из этих районов в северной Мексике плохо
исследованы. Промежуточной формой из Коауила было вероятно растение,
упомянутое Бакебергом и Килиан (1960), хотя все, что они сказали, были то,
что это растение происходил из «Nordmexiko», и его точное
местоположение не было известно. К сожалению, одна из их фотографий
(на странице 149) подписанная как Roseocactus intermedius —
почти точно A. kotschoubeyanus. Андерсон (1963) рассмотрел
R. intermedius nomen nudum и понизил его
до синонима A. fissuratus var. lloydii.
Однако, он не видел живого растения промежуточной формы до 1979 (EFA #5018, US,
DES). Мы оба теперь можем наблюдать эту промежуточную форму на нескольких
местообитаниях. Присутствие промежуточной формы между севером и югом в диапазоне почти
150 километров и между местобитаниями этих двух разновидностей,
говорит нам что A. fissuratus состоит из географических рас.
Мы заключаем, что таксон A. fissuratus характеризуется
непрерывно изменяющимися сосочками по форме и по конфигурации и имеет
переходные формы с севера на юг и что не стоит выделять части непрерывного как
внутривидовые таксоны. Нет никаких других характеристик достаточно отличающихся
в пределах вида, чтобы поддержать отделение на внутривидовом уровне.
Мы, также, были обеспокоены присутствием промежуточных форм
A. fissuratus от Куатро Синегас до Эстасион Марте в штате
Коауила. Кроме того, многие из этих районов в северной Мексике плохо
исследованы. Промежуточной формой из Коауила было вероятно растение,
упомянутое Бакебергом и Килиан (1960), хотя все, что они сказали, были то,
что это растение происходил из «Nordmexiko», и его точное
местоположение не было известно. К сожалению, одна из их фотографий
(на странице 149) подписанная как Roseocactus intermedius —
почти точно A. kotschoubeyanus. Андерсон (1963) рассмотрел
R. intermedius nomen nudum и понизил его
до синонима A. fissuratus var. lloydii.
Однако, он не видел живого растения промежуточной формы до 1979 (EFA #5018, US,
DES). Мы оба теперь можем наблюдать эту промежуточную форму на нескольких
местообитаниях. Присутствие промежуточной формы между севером и югом в диапазоне почти
150 километров и между местобитаниями этих двух разновидностей,
говорит нам что A. fissuratus состоит из географических рас.
Мы заключаем, что таксон A. fissuratus характеризуется
непрерывно изменяющимися сосочками по форме и по конфигурации и имеет
переходные формы с севера на юг и что не стоит выделять части непрерывного как
внутривидовые таксоны. Нет никаких других характеристик достаточно отличающихся
в пределах вида, чтобы поддержать отделение на внутривидовом уровне.
наверх
Орфография A. scapharostrus
Немного нужно остановиться на мнении (1991) Ханта, который верно
доказывает, что правильнее было бы называть этот таксон —
scaphirostris. Мы принимаем его коррекцию и называем этот таксон с
его поправкой написания повсюду в этой статье.
наверх
Состояние Ariocarpus (Anhalonium) elongatus Salm-Dyck
Андерсон (1964) причислял отдельный эпитет elongatus как синоним
A. retusus, типовым видам рода, который был описан
Шейдвейлером в 1858. Шейдвейлер не сохранил или выделил типовое растение,
так что Андерсон (1964) выделил неотип для A. retusus,
основанный на образце собранным им к востоку от города Сан Луис Потоси
(EFA #1200, POM; образец — также в DES). Х. Браво и
Санчез-Мехорада (1991) согласился с Андерсоном, что эпитет elongatus —
синоним A. retusus. Недавно Хант (1991) обсуждал спорный
статус отдельного эпитета elongatus, аккуратно суммируя различные
мнения по этому эпитету многочисленными авторами. Эпитет сначала использовался
Сальм-Диком (1850) в описании новой разновидности Anholonium,
в родовом имени созданным Лемайром в 1839. Как объяснено Андерсоном (1964),
род Anholonium является более поздним синонимом рода Ariocarpus,
и был опубликован для того же самого растения (A. retusus)
описанного Шейдвейлером годом ранее. К сожалению, Сальм-Дик не включил ни
иллюстрацию места из которого растение прибыло, ни описания цветка; любое
из них почти точно прояснил бы к какому растению оно относилось. И при этом
он не выделил или сохранил типового экземпляра. Проблема, как правильно
заявлено (1991) Хантом, что Сальм-Дик упоминал о сосочках A.
elongatus как о «напоминающих о листьях Crassula perfoliata
(описание подходящее для сосочков A. trigonus), принимая
во внимание, что Anhalonium prismaticum (теперь
Ariocarpus retusus) походил на листья Aloe retusa».
Ли (1937) верил, что таксон elongatus заслуживает статус отдельного
рода. Аналогично, Хант (1992) временно воспринимал его как отдельный вид.
Веттштейн (1933), с другой стороны, верил, что этот таксон вероятно относится
к A. trigonus, также как и Хант (1989). Хант (1991) далее
указал, что если бы это было истинно, то таксон теперь известный как
A. trigonus должен стать A. elongatus,
потому что это название было дано 40 годами ранее! Хааге (1977), в его
расширении «Kakteenlexikon» Бакеберга, сомневался, формулируя что
«этот вид или разновидность A. trigonus спорен».
Интересно, он продолжал говорить, что таксон был из Юмаве (Тамаулипас), хотя
это местоположение, конечно, не было заявлено Сальм-Диком в 1850. Как Хааге
мог знать оригинальное место произрастание растений, с которыми имел дело
Сальм-Дик? Если он был действительно правилен в заявлении, что таксон
elongatus происходил из Юмаве, то очень вероятно это —
тот же самый таксон, который в настоящее время известен как A.
trigonus.
Мы повторно исследовали эту проблему и в отсутствии описания
местоположения Сальм-Диком, поверим, что тождество таксона может быть
определено из другой части его описания A. elongatum.
Во втором параграфе описания Anhalonium elongatum он заявляет:
«apice pulvillo tomentoso instructa», что при переводе с Латинского
означает «вершина [сосочка] снабжена опушенной подушкой».
Мы верим, что он имел в виду маленькую подушечку на ареоле около кончика
сосочка, которая является разделительной характеристикой A. retusus.
Ariocarpus trigonus никогда не имеет такие подушечки в конце
ареол на его сосочках на взрослых растениях; они присутствуют только на
сосочках сеянцев.
 Дилемма относительно формы и длинны сосочков, описанных для
A. elongatum Сальм-Диком может быть разрешена,
исследованием отдельных растений A. retusus растущих около
примыкании дорог 57 и 80 вблизи Эль Хуизаче в штате Сан Луис Потоси.
Некоторые из этих растений, одно из которых Андерсон (1964) обозначил как
неотип A. elongatum (EFA #1077, POM), имеют особенно длинные,
конусобразные сосочки, которые достигают в длину до 5 сантиметров
(два дюйма) и с подобной формой как у A. trigonus.
Некоторый даже имеют плавный изгиб, такой же, как и у A. trigonus.
Однако, они отчетливо сине-зеленые по окраске, а не желто-зеленые как у
trigonus. Если популяцию в Эль Хуизаче экстенсивно исследовать,
то там найдутся растения с широко изменяющимися формами сосочка, в пределах
от 1:1 к 4:1 по отношению длинны к ширине. Фактически, A.
trigonus-подобные сосочки являются там исключением скорее,
чем правилом. Другими словами, эти популяция демонстрирует диапазон изменения
сосочка от «типичного» A. trigonus к имеющимся
у северных форм A. retusus. Популяция Эль Хуизаче не состоит
из двух видов растущих вместе, но — скорее популяция
A. retusus демонстрирует высокую вариабильность формы
сосочка. Кроме того, кончики сосочков, особенно в молодом возрасте, обычно имеют
подушечки на ареолах. Так, хотя Хант (1991) верил, что Андерсоновское (1964)
определение своего коллекционного растения #1077 как neotype A.
elongatus было «решительно ненадежно» мы чувствуем, что оно
является точным представителем неполного описания Сальм-Дика, насколько это
возможно и является представителем одного из многих вариантов A.
retusus.
Дилемма относительно формы и длинны сосочков, описанных для
A. elongatum Сальм-Диком может быть разрешена,
исследованием отдельных растений A. retusus растущих около
примыкании дорог 57 и 80 вблизи Эль Хуизаче в штате Сан Луис Потоси.
Некоторые из этих растений, одно из которых Андерсон (1964) обозначил как
неотип A. elongatum (EFA #1077, POM), имеют особенно длинные,
конусобразные сосочки, которые достигают в длину до 5 сантиметров
(два дюйма) и с подобной формой как у A. trigonus.
Некоторый даже имеют плавный изгиб, такой же, как и у A. trigonus.
Однако, они отчетливо сине-зеленые по окраске, а не желто-зеленые как у
trigonus. Если популяцию в Эль Хуизаче экстенсивно исследовать,
то там найдутся растения с широко изменяющимися формами сосочка, в пределах
от 1:1 к 4:1 по отношению длинны к ширине. Фактически, A.
trigonus-подобные сосочки являются там исключением скорее,
чем правилом. Другими словами, эти популяция демонстрирует диапазон изменения
сосочка от «типичного» A. trigonus к имеющимся
у северных форм A. retusus. Популяция Эль Хуизаче не состоит
из двух видов растущих вместе, но — скорее популяция
A. retusus демонстрирует высокую вариабильность формы
сосочка. Кроме того, кончики сосочков, особенно в молодом возрасте, обычно имеют
подушечки на ареолах. Так, хотя Хант (1991) верил, что Андерсоновское (1964)
определение своего коллекционного растения #1077 как neotype A.
elongatus было «решительно ненадежно» мы чувствуем, что оно
является точным представителем неполного описания Сальм-Дика, насколько это
возможно и является представителем одного из многих вариантов A.
retusus.
наверх
A. retusus — A. trigonus дилемма
Андерсон (1964), Х. Браво и Санчез-Мехорада (1991) и многие другие
восприняли A. retusus и A. trigonus как
отдельные виды. В 1994 мы посетили местообитание около Арамберри (Нуэво Леон),
с популяцией Ariocarpus, первооткрыватель идентифицировал их как
A. trigonus. Цветки некоторых растений из этой популяции
были описаны как красные, что явно разнится от имеющихся как у A.
retusus так и A. trigonus. Открытие этой популяции
Ariocarpus с «красными» цветками умножило вопросы относительно
обособленности этих двух таксонов, а также — не являются ли
эти растения новым таксоном. Растения из области Арамберри были исследованы
обоими из нас. В первое время, одни из нас (Андерсон) сомневался, какое это
растение — A. retusus или A.
trigonus. Популяция была не похожа ни точно на A. trigonus,
ни точно на A. retusus — единственные два
вида на которых они вообще похожи. Ниже приведенные характеристики, были
основанием для разделения A. retusus и A.
trigonus, но Арамберри и несколько других популяций показывает, что
имеются исключения или интерградации.
Характеристики:
Окраска Эпидермиса: У A. retusus обычно
серо-зеленый, но иногда более темно-зеленый без серого, как замечено в
популяциях в Сан Рафаэл и южнее Вилла Джуарез (S.L.P). Растения A.
trigonus обычно коричневато-зеленые до желто-зелёных. Эти две, обычно
раздельные, цветовые формы объединяются в популяции Ариокарпусов возле Арамберри.
Форма сосочка: Сосочки A. retusus имеют
форму от широких и коротких (1:1), с угловым килем до менее часто встречающихся
более тонких и длинных (1:4) без углового киля, последние найдены в Энтронкве
Хуизаче (S.L.P.), Сан Роберто (N.L) и других местах. Сосочки A.
trigonus имеют форму от тонких и длинных (1:4), к менее часто
встречающимся широким и коротким (1:1), никогда не имеют углового киля. В то
время как сосочки A. retusus обычно имеют треугольное
поперечное сечение с острыми кромками, таковые у A. trigonus
более или менее округлены. Однако, эти два типа граней встречаются вместе в
некоторых популяциях, таких как в Арамберри и Сан Рафаэл (S.L.P.).
Ареолы: Оба таксона имеют терминальные колючконесущие ареолы,
имеющиеся уже в стадии сеянца. Эти ареолы — обычно
присутствуют на сосочках зрелых растений A. retusus, но
отсутствуют у A. trigonus. Размещение этих ареольных
подушечек, которые наличествуют у A. retusus, изменяется от
значительно низкого до периферии кончика. Популяция Ариокарпусов в Арамберри
имеет отдельные растения несущие терминальные подушечки на ареолах и растения
совершенно без них. Даже незрелые сосочки на некоторых зрелых растениях не
имеют подушечки.
 Цветки: не имеется никаких значительных различий в
особенностях цветка между двумя таксонами, за исключением их окраски. Цветки
A. retusus были описаны как белые до розовато-белых и у
A. trigonus как светло-желтые или кремовые. Растения из
области Арамберри имеют цветки, окраска которых варьируется в диапазоне от
белого и кремово-белого с розовыми кончиками лепестков до гвоздично-темного
или фуксинового (того же самого окраса как и у A. fissuratus
или A. agavoides). В октябре 1995 одни из нас (Фитц Морис)
посетил одно из местобитания расположенное к северу от Арамберри, где были
изучены более чем 100 растений, 30 из них цвели. Цветки одного растения были
гвоздично-темной окраски, цветки двух растений имели розовые лепестки
околоцветника, в то время как все другие были белые со случайным оттенком
кремово-белого. Однако, эти растения, при обладании более или менее типичными
белыми цветками A. retusus, имели морфологические признаки
более напоминающие A. trigonus чем A. retusus.
Цветки: не имеется никаких значительных различий в
особенностях цветка между двумя таксонами, за исключением их окраски. Цветки
A. retusus были описаны как белые до розовато-белых и у
A. trigonus как светло-желтые или кремовые. Растения из
области Арамберри имеют цветки, окраска которых варьируется в диапазоне от
белого и кремово-белого с розовыми кончиками лепестков до гвоздично-темного
или фуксинового (того же самого окраса как и у A. fissuratus
или A. agavoides). В октябре 1995 одни из нас (Фитц Морис)
посетил одно из местобитания расположенное к северу от Арамберри, где были
изучены более чем 100 растений, 30 из них цвели. Цветки одного растения были
гвоздично-темной окраски, цветки двух растений имели розовые лепестки
околоцветника, в то время как все другие были белые со случайным оттенком
кремово-белого. Однако, эти растения, при обладании более или менее типичными
белыми цветками A. retusus, имели морфологические признаки
более напоминающие A. trigonus чем A. retusus.
 То есть многие из этих растений имели сосочки изгибающиеся к центру, с
отношение «длинны к ширине» 3:1 или большим, тёмно-окрашенные
сосочки с закругленными краями и они совершенно не имели ареольной подушечки.
Другие растения казались явными промежуточными звеньями, с сосочками по форме
напоминающими A. retusus, но желто-зеленой окраски, как у
A. trigonus и с отсутствием на их кончиках околюченных
ареольных подушечек. Во второй области к северу от Арамберри имелось более
100 цветущих растений. Больше чем 95% цветков этих растений были белыми или
почти белыми, у остальных окраска варьировалась в пределах от цветков с
розовыми кончиками лепестков до полностью темно-розовых. Снова, многие из
растений, казались морфологически более близкими к A. trigonus
нежели чем к A. retusus. В третьей посещенной области, к
западу от Арамберри, имелись несколько сотен растений, приблизительно 30%
из которых цвели. Приблизительно 99% этих цветков были розовыми до
темно-розового по окраске, в полную противоположность тем двум
популяциям на севере. Важно подчеркнуть,
что все популяции около Арамберри имеют экземпляры с формами сосочка,
изменяющимися от типичного для A. retusus до типичного для
A. trigonus, но эти вариации не всегда коррелируются с
окраской цветка. Случайные промежуточные формы между A. retusus
и A. trigonus наблюдаются и в культуре. Такие промежуточные
формы также наблюдались в нескольких местах, преимущественно в области
Арамберри, но также и в Сан Луис Потоси у Эль Хуизаче и у Сан Рафаэль.
Мы думаем, что имеются и другие популяции с промежуточными формами.
То есть многие из этих растений имели сосочки изгибающиеся к центру, с
отношение «длинны к ширине» 3:1 или большим, тёмно-окрашенные
сосочки с закругленными краями и они совершенно не имели ареольной подушечки.
Другие растения казались явными промежуточными звеньями, с сосочками по форме
напоминающими A. retusus, но желто-зеленой окраски, как у
A. trigonus и с отсутствием на их кончиках околюченных
ареольных подушечек. Во второй области к северу от Арамберри имелось более
100 цветущих растений. Больше чем 95% цветков этих растений были белыми или
почти белыми, у остальных окраска варьировалась в пределах от цветков с
розовыми кончиками лепестков до полностью темно-розовых. Снова, многие из
растений, казались морфологически более близкими к A. trigonus
нежели чем к A. retusus. В третьей посещенной области, к
западу от Арамберри, имелись несколько сотен растений, приблизительно 30%
из которых цвели. Приблизительно 99% этих цветков были розовыми до
темно-розового по окраске, в полную противоположность тем двум
популяциям на севере. Важно подчеркнуть,
что все популяции около Арамберри имеют экземпляры с формами сосочка,
изменяющимися от типичного для A. retusus до типичного для
A. trigonus, но эти вариации не всегда коррелируются с
окраской цветка. Случайные промежуточные формы между A. retusus
и A. trigonus наблюдаются и в культуре. Такие промежуточные
формы также наблюдались в нескольких местах, преимущественно в области
Арамберри, но также и в Сан Луис Потоси у Эль Хуизаче и у Сан Рафаэль.
Мы думаем, что имеются и другие популяции с промежуточными формами.
Семена: семена A. retusus и A.
trigonus схожи в целом по всем аспектам и могут быть микроскопически
дифференцированы от семян других разновидностей Ariocarpus.
Географическое распределение: A. retusus
широко распространён повсюду в пустыне Чихуахуа в северной Мексике на высоте
более чем 1000 метров, с самым высоким известным месторождением более чем в
2000 метрах около Сан Луис Потоси. Ariocarpus trigonus происходит
прежде всего из зоны околюченной растительности на высотах ниже 1000 метров.
Одно из известных месторождений A. trigonus находится к
северо-западу от Палмиллас, Тамаулипас, находится на высоте в 1400 метров
(Glass & Foster #665), и он встречается в зоне измененной растительности
пустыни Чихуахуа в долине Джаумаве на высотах меньше чем 1000 метров. Около
Ллера, Тамаулипас, он встречается на высоте в 100 метров. Таким образом,
вообще имеется два географически отделенных таксона.
(Карта 3), показывает
географическое размещение известных и собранных авторами —
A. retusus и A. trigonus. Однако популяции
около Арамберри отстоит только на 15-20 километров (9-12.5 миль) от типичного
местообитания A. trigonus и маленького, но точно не
известного расстояния от того, что считается областью произрастания для
A. retusus. Другие области произрастания, такие как у Вилла
Джуарез, Сан Рафаэль, и Энтронкве Хуизаче, также показывают объединение
характеристик.
Имеются три возможных выбора для имеющих дело с A.
retusus — A. trigonus дилеммой.
Первый выбор это принять A. retusus и A.
trigonus как представителей крайних членов непрерывной линии, по
подобию виденного у A. fissuratus и у северно-южных
переходных форм A. retusus, как описано в более ранней
статье (Андерсон, 1964). Однако, в большинстве их ареалов A. retusus
и A. trigonus остаются различными. Если, действительно переходные
формы существуют, то все популяции должны признаться как A. retusus,
и имя A. trigonus должен быть понижено до синонима.
Рассматривая популяцию в Арамберри, мы не верим, что это лучшее всего
характеризует ситуацию.
Второй выбор — продолжить признавать A. retusus
и A. trigonus как отдельные виды, несмотря на свидетельство их
гибридности в Арамберри
(Карта 3).
Случайные промежуточные формы появляются в садоводствах (Хирао, 1979), но,
за исключением совокупностей Арамберри, фактически невозможно спутать два
таксона в поле. Два таксона, как отмечено выше, должны иметь определенные
наборы характеристик, которые включают различные типы развития ареол. Изучения
гибридизации Луксом и Стаником (1983) и наблюдения над многочисленными сосочками
от собранных в поле растений поддерживают гипотезу, что это может быть
популяция, совместно использующая гены обоих видов. Кроме того, популяция в
Арамберри отделена от мест произрастания A. trigonus только
приблизительно 20 километрами (12.5 миль) и имеется каньон сквозь
Сьерра в той точке, которая могла облегчить генетический обмен. Известно, что
никакие другие виды Ariocarpus не гибридизируются так просто
(на месте); при этом фактически, некоторые виды даже растут вместе. Поэтому,
вероятно неуместно дальше продолжать признавать эти два таксона как различные
виды.
Третий выбор это признать A. trigonus как
внутривидовой таксон A. retusus, отражая факт, что два
таксона могут скрещиваться. Так A. trigonus был бы подвидовым
таксоном высоко изменчивого вида A. retusus, с дифференцированным
географическим размещением, частично перекрывающимся, но растущим на более
низких высотах, у него отсутствуют несущие колючки ареольных подушечек и для
него характерны цветки с желтым или кремообразно-белым окрасом. Растения
Арамберри, таким образом, являются гибридной популяцией в ряде переходных форм
протянувшихся с запада на восток, между типичными для пустыни Чихуахуан формами
A. retusus и его подвида или разновидности trigonus,
обычно растущего на более низких высотах в зонах околюченной растительности.
Итак, дилемму A. retusus — A.
trigonus не легко разрешить. То, что замечено в Арамберри это вероятно
соединение двух наборов характеристик. Если это не больше, чем соединение
географических рас, то эта дилемма заключается в том, что признать ли формально
критическую форму (типичный A. trigonus) как внутривидовой
таксон. Мы уже обсудили, что род Ariocarpus содержат сильно изменчивые
таксоны. Это было показано в ранней статье Андерсона (1964) на примере
вариабельности сосочков A. retusus с севера на юг. В этой
статье мы говорили, что аналогично ¡ A. fissuratus
настолько изменчив, и что он не должно быть подразделен на внутривидовые таксоны.
Мы заключаем, что A. retusus и A.
trigonus являются еще одним образцом большой изменчивости и что они
не должны быть сохранены как отдельные виды. Итак, мы предлагаем, чтобы A.
trigonus был понижен к внутривидовому таксону подвида: Ariocarpus
retusus Scheidweiler subsp. trigonus (F.A.C. Weber)
Anderson and Fitz Maurice, comb. et stat nov. (Basionym = Anhalonium
trigonum F.A.C. Weber, Dict. Hort. Bois 90. 1893).
наверх
Статусы сохранности Ариокарпусов
Все виды Ariocarpus включены в Приложение 1 CITES
— в результате акции предпринятой на встрече CITIES в Киото,
Япония, в 1992. Однако это не подразумевает, что все эти виды имеют статус
«Уязвимый» или «В опасности». Одно из оправданий для
включения всех видов в Приложении 1 было то, что более обычные
виды могут быть легко перепутаны с наиболее редкими. Нижеследующее подводит
итог наших наблюдений относительно состояния сохранения видов Ariocarpus.
Наши категории угрозы соответствуют новой системе «Species Survival
Commission» IUCN (1994).
 A. agavoides
(Карта 3) —
этот вид один из наиболее редких в его известной области распространения и его
количество продолжает сокращаться разрушением местообитания и его запрещенным
сбором. Эрнандез-Барера (1992) проводил демографический подсчёт в известном
месторождении A. agavoides и обнаружил несколько сотен
растений. Недавно было обнаружено новое местообитание этого вида,
располагающееся на значительном удалении от типового. Необходима дальнейшая
полевая работа, что бы определить область распространения этого вида, но мы
чувствуем, что это придает ему соответствующий статус «В опасности» (EN).
A. agavoides
(Карта 3) —
этот вид один из наиболее редких в его известной области распространения и его
количество продолжает сокращаться разрушением местообитания и его запрещенным
сбором. Эрнандез-Барера (1992) проводил демографический подсчёт в известном
месторождении A. agavoides и обнаружил несколько сотен
растений. Недавно было обнаружено новое местообитание этого вида,
располагающееся на значительном удалении от типового. Необходима дальнейшая
полевая работа, что бы определить область распространения этого вида, но мы
чувствуем, что это придает ему соответствующий статус «В опасности» (EN).
 A. bravoanus
(Карта 1) —
этот новый вид, с его разновидностью hintonii, в настоящее время
известен встречающимся только в трех ограниченных местах. Одно место —
очень близко к деревне, и имеется серьезная угроза разрушения местообитания.
Это место также известно и собирателям и имеется несомненные свидетельства их
действий. Известно, что растения этой разновидности были конфискованы в Европе.
Другие два местообитания далеки от деревень или других человеческих действий,
и поэтому имеется меньшая вероятность, что в тех местах будет происходить
разрушение местообитаний. Однако, число индивидуальных растений в каждом
месторождении — исчисляется только в тысячах, так что
собиратели могут быстро разорить любую из этих малых популяций. Фактически,
мы недавно посетили единственное известное местообитание A. bravoanus
subsp. bravoanus. и обнаружили свидетельства запрещенного сбора.
Местные жители также используют несколько видов Ariocarpus, включая
A. bravoanus, для лечебных целей. Необходима дальнейшая
полевая работа для определения — встречается ли эта
разновидность в других местах. В настоящее время мы считаем, что этот вид
должен рассматриваться под статусом «В опасности» (EN) из-за его
очень ограниченного распространения и лёгкой уязвимости.
A. bravoanus
(Карта 1) —
этот новый вид, с его разновидностью hintonii, в настоящее время
известен встречающимся только в трех ограниченных местах. Одно место —
очень близко к деревне, и имеется серьезная угроза разрушения местообитания.
Это место также известно и собирателям и имеется несомненные свидетельства их
действий. Известно, что растения этой разновидности были конфискованы в Европе.
Другие два местообитания далеки от деревень или других человеческих действий,
и поэтому имеется меньшая вероятность, что в тех местах будет происходить
разрушение местообитаний. Однако, число индивидуальных растений в каждом
месторождении — исчисляется только в тысячах, так что
собиратели могут быстро разорить любую из этих малых популяций. Фактически,
мы недавно посетили единственное известное местообитание A. bravoanus
subsp. bravoanus. и обнаружили свидетельства запрещенного сбора.
Местные жители также используют несколько видов Ariocarpus, включая
A. bravoanus, для лечебных целей. Необходима дальнейшая
полевая работа для определения — встречается ли эта
разновидность в других местах. В настоящее время мы считаем, что этот вид
должен рассматриваться под статусом «В опасности» (EN) из-за его
очень ограниченного распространения и лёгкой уязвимости.
A. fissuratus
(Карта 1) —
один из наиболее широко распространенных видов рода, имеются немногие
свидетельства влияния на него людей. Некоторое разрушение местобитаний может
воздействовать на местные популяции, такие как в местах около Паррас, но,
вообще, эта разновидность соответственно должна рассматриваться как обладающая
статусом «Низкий риск» (LR).
 A. kotschoubeyanus,
(Карта 2) —
другой относительно обычный вид с широким распространением, этот
таксон — вероятно вне опасности. Тем не менее,
сельскохозяйственная деятельность встречается на многих илистых (наносных)
равнинах, где A. kotschoubeyanus обычно растет, так что
местные популяции будут, вероятнее всего ужиматься. Мы полагаем, что этот вид
должен обладать статусом «Уязвимый» (VU).
A. kotschoubeyanus,
(Карта 2) —
другой относительно обычный вид с широким распространением, этот
таксон — вероятно вне опасности. Тем не менее,
сельскохозяйственная деятельность встречается на многих илистых (наносных)
равнинах, где A. kotschoubeyanus обычно растет, так что
местные популяции будут, вероятнее всего ужиматься. Мы полагаем, что этот вид
должен обладать статусом «Уязвимый» (VU).
 A. retusus,
(Карта 3) —
этот вид является, как и обычным, так и широко распространенным повсюду в
пустыне Чихуахуан. Не имеется фактически никакой опасности, что он будет
уничтожен или подвергнется неблагоприятным воздействиям людскими стараниями,
тем не менее, некоторые местные популяции могли быть уничтожены разрушением
местообитаний. Многие из местобитаний подвида trigonus были изменены
для ведения сельскохозяйственной деятельности. Но это всё ещё компенсируется
довольно широким его распространением, как к востоку от Сиерра Мадре Ориентал,
так и в пределах долины Джаумаве. Местные популяции были разрушены
человеческими действиями, но Мартинез и другие (1993), показали что, растение
изобилует во многих областях долины Джаумаве. Этот вид должна рассматриваться
как имеющий статус «Низкий риск» (LR).
A. retusus,
(Карта 3) —
этот вид является, как и обычным, так и широко распространенным повсюду в
пустыне Чихуахуан. Не имеется фактически никакой опасности, что он будет
уничтожен или подвергнется неблагоприятным воздействиям людскими стараниями,
тем не менее, некоторые местные популяции могли быть уничтожены разрушением
местообитаний. Многие из местобитаний подвида trigonus были изменены
для ведения сельскохозяйственной деятельности. Но это всё ещё компенсируется
довольно широким его распространением, как к востоку от Сиерра Мадре Ориентал,
так и в пределах долины Джаумаве. Местные популяции были разрушены
человеческими действиями, но Мартинез и другие (1993), показали что, растение
изобилует во многих областях долины Джаумаве. Этот вид должна рассматриваться
как имеющий статус «Низкий риск» (LR).
 A. scaphirostris, (Карта 2) —
хотя этот вид, как известно, происходит только из одной долины в штате Нуэво
Леон, единственно возможная угроза его существованию это незаконный сбор этого
растения. Его типичное местообитание не соответствует любым человеческим
действиям, включая сельское хозяйство. Всё же, из-за его ограниченного
распространения мы рассматриваем этот вид, как имеющий статус
«Уязвимый» (VU).
A. scaphirostris, (Карта 2) —
хотя этот вид, как известно, происходит только из одной долины в штате Нуэво
Леон, единственно возможная угроза его существованию это незаконный сбор этого
растения. Его типичное местообитание не соответствует любым человеческим
действиям, включая сельское хозяйство. Всё же, из-за его ограниченного
распространения мы рассматриваем этот вид, как имеющий статус
«Уязвимый» (VU).
наверх
Ключ к определению видов и подвидов рода Ariocarpus
 |
|
A |
|
Зрелые растения обычно не превышают 9 см в диаметре |
|
|
|
| |
B |
|
Сосочки разделены, не плотно собранны у основания |
|
|
| |
|
C |
|
Длина сосочков по крайней мере вдвое превышает ширину, поверхность их не
бородавчатая. Не из Сан Луис Потоси |
|
|
| |
|
|
D |
|
Сосочки
несут околюченные ареольные подушечки. Тамаулипас —
1. A. agavoides |
|
|
| |
|
|
DD |
Сосочки не несут
околюченные ареольные подушечки, Нуэво Леон —
2. A. scaphirostris |
|
|
| |
|
CC |
Длина сосочков менее чем вдвое превосходит ширину,
поверхность их бородавчатая. Сан Луис Потоси —
3. A. bravoanus |
|
|
| |
|
|
E |
|
Сосочки отчётливо приподняты, ареольная бороздка
отсутствует — subsp. bravoanus |
|
|
| |
|
|
EE |
Сосочки не приподняты отчётливо, с ареолой в виде
бороздки — subsp. hintonii |
|
|
| |
BB |
Сосочки не разделены, плотно собраны у основания
— 4. A. kotschoubeyanus |
|
|
| AA |
Зрелые растения, обычно больше 9 см в диаметре |
|
|
| |
B |
|
Сосочки с ареольной бороздкой,
бородавчатые, закруглённые на конце — 5. A.
fissuratus |
|
|
| |
BB |
Сосочки без арельной бороздки,
не бородавчатые, заострённые на конце — 6. A.
retusus |
|
|
| |
|
C |
|
Сосочки серые —
или голубовато-зелёные, обычно несут околюченную ареолу, расположенную вблизи
кончика, концы сосочков не изгибаются, цветки белые до розовато-белых
— subsp. retusus |
|
|
| |
|
CC |
Сосочки желтовато-зелённые,
никогда не имеют околюченной ареолы на кончиках, изогнуты на конце,
цветки жёлтые до кремово белых —
subsp. trigonus |
|
|
наверх
Благодарности
We wish to thank the following for providing facilities and financial
support for this project: Cante A.C., Desert Botanical Garden, World Wildlife
Fund, and the Cactus and Succulent Society of America Research Fund. We also
thank Dr. Hector Hernandez for making scanning electron micrographs of the
seeds of several species of Ariocarpus. George Hinton developed the computer
software for generating the maps; his expertise and advice is much appreciated.
A portion of this study was conducted under the direction of Cante A.C., San
Miguel de Allende, Gto., with the authorization of La Direccion General
de Aprovechamiento Ecologico de los Recursos Naturales del Institute Nacional
de Ecologica, SEDESOL, oficios numero 01516 y 05860. We also wish to thank the
Mexican government for issuing collecting permit #DO0 (2) 2532 to allow us to
collect a small number of Ariocarpus plants for study, especially during
flowering season. The plants are presently growing at Cante A.C. and the
Desert Botanical Garden.
наверх
Литература
Anderson, E.F. 1958. A recent field trip in search of Ariocarpus
Cact. Succ. J. (U.S.): 30:171-174.
Anderson, E.F. 1960. A revision of Ariocarpus (Cactaceae)
I The status of the proposed genus Roseocactus
Amer. J. Bot. 47(7):582-589.
Anderson, E.F. 1962. A revision of Ariocarpus (Cactaceae)
II The status of the proposed genus Neogomesia
Amer. J. Bot. 49(6):615-622.
Anderson, E.F. 1963. A revision of Ariocarpus (Cactaceae)
III Formal taxonomy of the subgenus Roseocactus
Amer. J. Bot. 50(7):724-732.
Anderson, E.F. 1964. A revision of Ariocarpus (Cactaceae)
IV Formal taxonomy of the subgenus Ariocarpus
Amer. J. Bot. 51(2):144-151.
Backeberg, C. 1966. Kakteenlexikon Veb Gustav Fischer, Jena.
Barthlott, W. 1979. Cacti Stanley Thornes, Cheltenham.
Berger, A. 1925. Roseocactus, a new genus of Cactaceae J.
Wash.(D.C.) Acad. Sci. 15:43-48.
Bravo H.,H., and H. Sanchez-Mejorada 1991. Las Cactaceas de Mexico
Vol 2, p. 260. Universidad Nacional Autonoma de Mexico, Mexico City.
Fitz Maurice, W.A., and T. E. Davis. 1987. Fieldnotes Cact.
Succ. J. (U.S.): 59:144-145.
Haage, W. 1977. Curt Backeberg's Cactus Lexicon Trans. of 3rd ed.
Blandford Press, Poole.
Hernandez, H., and E.F. Anderson. 1992. A new species of
Ariocarpus (Cactaceae) Bradleya 10:1-4.
Hernandez-Barrera, J.T. 1992. Aspectos biologicos de Ariocarpus
agavoides (Castaneda) Anderson. Cact. Suc. Mex. 37:40-45.
Hirao, H. Color encyclopedia of cacti .Tokyo: Seibundo Shinkosha PubI. Co.,Ltd.
Hunt, D. 1989. Cactaceae. In S.M. Walters et al., eds. The European Garden
Flora. Vol. 3, p 277. Cambridge University Press.
Hunt, D.1991. Notes on miscellaneous genera of Cactaceae (with N.P. Taylor).
Bradleya 9:83.
Hunt, D.1992. CITES Cactaceae checklist. Royal Botanic Gardens, Kew.
IUCN. 1994 IUCN. Red List categories. IUCN. Gland.
Lee. M.H. 1937. Ariocarpus Cact. Succ. J. (U.S.): 9:71-77.
Lemaire, C. 1839. Cactearum genera nova et species nova en horto Monville.
Paris.
Lux, A., and R. Stanik. 1983. Hybridisation in der Gattung Ariocarpus
Scheidweiler Kakt. Sukk. 34:267-271.
Marshall, W.T. 1946. Revision of the genus Ariocarpus Cact. Succ.
J. (U.S.): 18:55-56.
Martinez, J.G., H. Suzan A., and C.A. Salazar O. 1993. Aspectos ecologicos y
demograflcos de Ariocarpus trigonus (Weber) Schumann. Cact. Suc. Mex
38:30-38.
Salm-Dyck, J. de. 1850. Cacteae in horto Dyckensi cultae. Anno 1849. Apud Henry
& Cohen, Bonn.
Scheidweiler, M.J. 1838. Descriptio diagnostica nonnullarum Cactearum quae a
domino Galeotti in provinciis Potosi et Guanaxato regni Mexicani inveniuntur.
Genus Ariocarpus. Bull. Acad. Sci. Brux. 5:491-492.
Schumann, K. 1898. Gesamtbeschreibung der Kakteen. J. Neumann, Neudamm.
Stuppy, W., and N.P. Taylor. 1989. A new variety of Ariocarpus
fissuratus (Cactaceae). Bradleya 7:84-88.
Wettstein, R. von. 1933. Die Bildung von Assimilationsorganen bei Kakteen
und die Erscheinung der Irreversibilitat. Kakteenkunde 1933:11-13.
Перевод А. Ю. Никитина 2001 г. |
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);)
;void(0);){kind=link}
;void(0);){kind=link}
;void(0);){kind=link}