Опыление Стапелиевых.
GERALD S. BARAD, M.D. Email: gbarad@eclipse.net
CSJ (U.S.), Vol. 62, 130-140.
В течение последних десяти лет, я разрабатывал методы искусственного
опыления стапелиевых. Эта работа проводилась в моей большой коллекции растений
находящейся в Флемингтоне, штат Нью-Джерси. Мне пришлось преодолеть множество препятствий
из-за того, что растения находились вне привычных им условий пустыни.
К сожалению, некоторые проблемы по контролю за здоровьем растений остались нерешёнными
и как результат, потеря подопытных растений, которые были уже
успешно опылены. Мне кажется, что к настоящему времени я приобрёл достаточно опыта
для того, что бы поделиться моими наблюдениями и теми методами, которые
я использую сейчас.
В каждый сезон цветения стапелиевых мною
производились приблизительно тысяча опылений. Хотя у моих растений
время цветения разбросано на весь год, но пик его в этом регионе
приходится на время между поздней весной и поздней осенью. Некоторые
виды, такие как Piaranthus, по-видимому, ограничивают своё цветение
только лишь осеннним периодом, в то время, как другие открывают свои
цветки нерегулярно во время всего возможного периода цветения.
У стапелиевых имеется большое
число вариантов размеров и форм цветков, изменяющихся в пределах от
огромных цветков Stapelia gigantea до крошечных цветков у
Echidnopsis и Rhytidocaulon. Все цветки стапелиевых демонстрируют
различные отличия от основной модели которая была хорошо описана
используя Orbea variegata как образец.
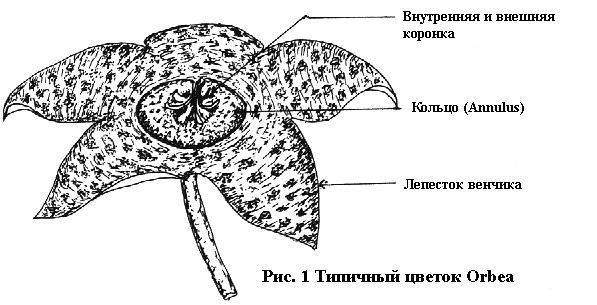
На рисунке 1, представлен характерный пяти лепестковый
цветок. По случайности, может появиться ненормальный (причудливый) цветок с четырёх или
шестью лепестковой структурой. В редчайших случаях появляются семи
лепестковые или полу-махровые цветки. Я однажды
наблюдал пример трёх лепесткового цветка у Duvalia sulcata который
вероятно был шести лепестковым, но со слившимися соседними лепестками.
Трёх лепестковые цветки Piaranthus (иллюстр. № 3) также наблюдались, и
недавно, был отмечен двух лепестковый у гибридной Huernia. Лепестки
венчика (corolla lobes) в основном кожистые или
полу-суккулентные. Их окраска изменяется внутри различных оттенков
коричневого, красного и жёлтого с отличающимися рисунками — от чисто
коричневой у Stapelia gigantea, до сильно орнаментированной, подобно
персидскому ковру, как у Edithcolea grandis. Считается, что большая
часть их опылителей это мухи или гнус, которые привлекаются окраской и
запахом, напоминающим запах гниющего мяса, навозной жижи или другой
падали и мерзости. Один из южно-аравийских видов, Caralluma
plicatiloba, имеет запах напоминающий запах от стойла верблюда в
нашем местном зоопарке. В редких случаях мы можем наблюдать
светло-розовые, белые или зелённые цветки, некоторые из них имеют
отчётливый и приятный запах пчелиного воска. Цветки с таким запахом,
такие как у Stapelia flavopurpurea, вероятно могут опыляться
пчёлами.
Правильное восприятие техники искусственного
опыления требует понимания основ строения стапелиевых цветков и
биологии их опыления. В отличие от цветков большинства высших
растений, которые опыляются переносом крупинок зернистой пыльцы с
пыльника на пестик, опыление у стапелиевых включает в себя высоко
специализированную структуру — поллинарий (pollinarium).
Для правильного обсуждения
поллинария нужно принять значения терминов используемых в описании
его частей и всех остальных органов у цветков стапелиевых. Кажется,
что ныне учёные пришли к общему соглашению, касающемуся названий
главных частей цветка (см. Рис № 1). Но с другой стороны, одни и те
же части поллинария имеют различные названия. Ньютон попытался
разрешить некоторые из этих различий путём использования термина
«биполлинарий (bipollinarium)». Но, кажется, этот термин так и не
будет признан. В его работе также рассматривается некоторое кол-во
более ранних публикаций по терминологии поллинария. Так как
поллинарий это структура, с которой большая часть этой статьи имеет
дело, необходимо выбрать понятную группу названий. Работа Букман
(Bookman) представляется мне, как широко признанная среди пишущих на
тему суккулентных ластовневых. Она быть может и не подходит,
поскольку её статья имеет дело с Asclepias speciosa, несуккулентным
ластовниевым у которого механизм введения поллиния в чём-то
отличается от того же у стапелиевых. Её статья также включает
обширный обзор предшествующих публикаций по теме терминологии
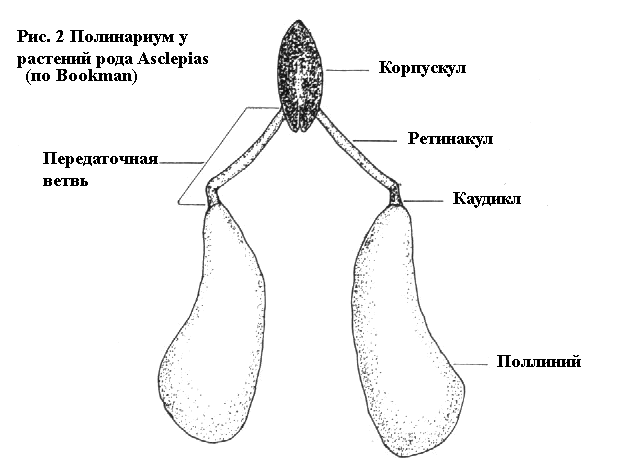
поллинария. Согласно Букман, и как мы видим на Рисунке 2, поллинарий
состоит из двух пыльцовых мешочков (pollen sacs) — поллиниев
(pollinia), каждый из них присоединён к каудиклу (caudicle) который
в свою очередь с изгибом
прикрепляется к ретинакулу (retinaculum). Каудикл и ретинакул
составляют каждый свою часть передаточной ветви (translator arm).
Передаточные ветви с каждой стороны прикрепляются к центральной
структуре — корпускулу (corpusculum). Корпускул и обе передаточных
ветви вместе называются трансляционным аппаратом.
Очень обширный отчёт Шилл и Джэкил (Schill, Jakel)
касается 114 родов ластовневых, при его подготовке использовались
как сканирующий электронный микроскоп, так и световой микроскоп. Они
тоже используют термин поллинарий для всей целиком опыляющей
структуры. В их статье, центральная структура называется
«klemmkörper», что можно перевести как «корпускул
(тельце?)(corpuscle)». Структуры присоединяющие его к поллиниям
называются каудиклами. Они описывают четыре основных типа каудиклов
которые были ими названы: односоставной нитевидный, двусоставной
нитевидный, крылатый и плоский.
Шнепф, Витциг и Шилл (Schnepf,
Witzig, Schill) показали, используя электроннный микроскоп, что
трансляционный аппарат сформирован как продукт секреции нижележащих клеток.
Целиком эта структура, поллинарий,
состоит из трансляционного аппарата и двух прикреплённых поллиниев.
Так как терминология Букман базировалась на Asclepias, необходимо
привнести некоторые интерпретации, что бы использовать те же самые
термины применимо к несколько отличающимся поллинариям стапелиевых.
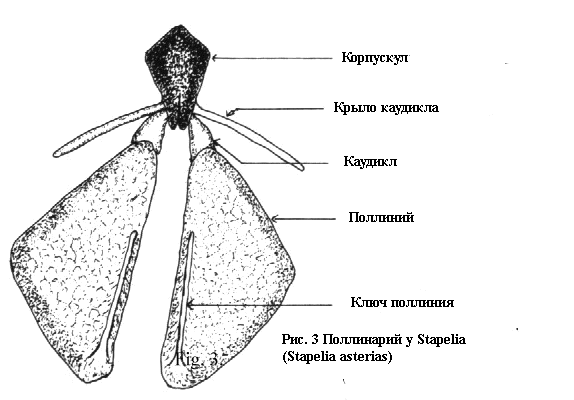
На Рисунке 3 представлен типичный стапелиевый поллинарий. Необходимо
отметить, что каудикл прикрепляющий поллиний к остальной части
трансляционнного аппарата, не присоединяется к концу той части,
которая обозначается как ретинакул у Asclepias. У стапелиевых
наблюдается значительное изменение в способе, которым присоединяется
каудикл к остальной части трансляционного аппарата. В
некоторых случаях прикрепление происходит в средней части крыла
каудикла, в то время как в других он присоединяется непосредственно
к корпускулу. Имеются также значительные изменения в длине и ширине
крыла каудикла. По всей видимости, будет лучше, если мы ограничим
использование термина ретинакул, к тем видам ластовневых у которых
структура его напоминает структуру Asclepias. У стапелиевых эту
структуру, присоединённую к корпускулу лучше именовать — крыло
каудикла и каудикл. Два каудикла и корпускул принято рассматривать
как трансляционный аппарат.
Из работы Букман, совершенно не ясно, полагает ли
она, что пыльцовые трубки (pollen tubes) появляются из вогнутой или
из выпуклой стороны поллиния. В своём тексте
она заявляет, что «пыльцовые трубки появляются из более изогнутого
края» и затем описывает обнаружение другим автором выемок в стенке
выпуклого края там, где
пыльцовые трубочки сперва появляются. Её диаграмма показывает
пыльцовые трубочки появляющиеся из вогнутого края. Чатурведи
(Chaturvedi) демонстрирует прорастание поллиния Cynanchium «in
vitro», которое происходит на краю удалённом от центра поллинария.
Это напоминает такую же
структуру у Asclepias, у которых пыльцовые трубки появляются из их
внешнего, вогнутого края. Но у поллиния Asclepias отсутствует
структура, которая специфически адаптирована для прикрепления к
принимающей (receptive — восприимчивой) области после того, как у
этого типа поллиния вся структура целиком вдвигается в принимающую
область которая описывается как щель (просвет, отверстие) между
крыльями пыльника (the anther wings).
У стапелиевых наблюдается
другой механизм прикрепления поллиния. Поллинии стапелиевых, которые
рассматриваются «in situ», имеют небольшие стержнеобразные
структуры, обширно присоединённые к внутренней, более
вышестоящей поверхности поллиния. Эти структуры называются
якорными каймами (anchor margin) или волнистым гребнем (wave crest)
у других авторов. Во время опыления, такая стержнеобразная структура
вдвигается во внутрь принимающей области подобно ключу и замку. По
этой причине, я предпочитаю, упоминать о них, как о ключах поллиния.
Это можно увидеть на Рисунке 3. Принимающая часть изменяется по
форме у различных родов стапелиевых, но всегда следует за общей
моделью паза, который открыт по направлению к основанию цветка и
сужается к вершине. У Huernia и Duvalia этот паз сравнительно
широкий, в то время как у большинства других родов он узок. Другие
авторы ссылаются на этот паз, как на направляющие поручни или
крылья пыльника. Для того, что бы сохранить функциональные аналогии
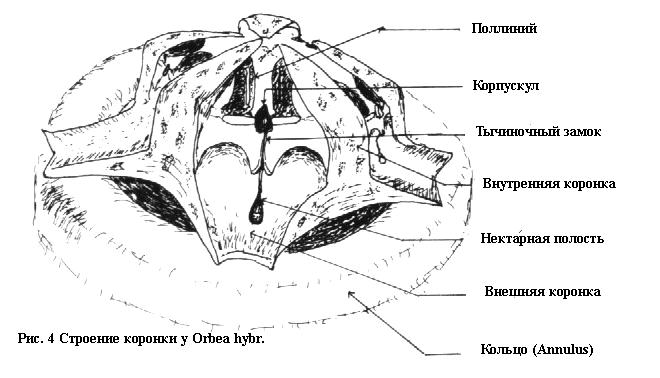
с ключом и замком, я предпочту называть этот паз тычиночным замком
(staminal lock) (Смотри рис. 4) Для упрощения, в остальной части этой
статьи я буду ссылаться на эти структуры, как на ключ и замок.
Таким образом, мы можем видеть, что стапелиевые растения отличаются
от Asclepias в основном только по наличию у них ключа вставляемого
во внутрь замка, нежели чем по наличию целостного поллиния
вставляемого между крыльями пыльника.
Я теперь возвращаюсь к методам удаления поллинария и прикрепления
поллинария. Эти растения всецело зависят от
насекомых опылителей для того, что бы завершить свой естественный процесс
опыления. Большинство из них опыляется или мухами питающимися
падалью или мелким гнусом, которые бывают привлечены
расцветкой напоминающей падаль и отвратительным запахом. Когда
насекомое садится на цветок, оно пытается достичь нектара или же
поместить свой яйцеклад на цветок, что бы отложить яйца. В процессе
выполнения этого, часть насекомого часто становится пойманной в
замке и в процессе само-высвобождения пленённая часть подтягивается
вверх по направлению бороздки в корпускуле, которая является
продолжением желобчатого замка. Насекомое всё так же продолжает свои
попытки освободиться и делая это оно отрывает целиком
поллинарий от его места прикрепления. Фотографии #1 и #2 показывают
муху пойманную корпускулом Stapelia flavopurpurea. В этом случае,
муха была поймана, но она была не достаточна сильна для того, что бы
удалить поллинарий.
После наблюдений за некоторым числом пойманных мух,
я заметил, что структура которая обычно попадает в такую западню,
это маленькая щетинка на ноге или хоботке. Целая нога или
хоботок представляют слишком большой объект для проникновения в
узкую щель замка и корпускула. Возможно, у более мелкого гнуса может
происходить и такое. Я отметил при искусственном опылении, что
поверхность поллиния, которая находится вдали от его прикрепления
перед удалением является той, которая должна быть обращённой к
замку, если введение должно быть завершено (выполнено). Это истинно
из-за угла наклона ключа на ту сторону. Я полагаю, что имеется
потребность в этой ориентации, которая объясняет функции внутренней
и внешней коронок (outer and inner coronas). Эти структуры нужны
для ограничения направления
входа опылителей, так что бы поллинарий был удалён и затем, что бы
сориентировать насекомое способом который способствует введению во
время посещения насекомым следующего цветка. Это кажется более
критичным у одних родов, нежели чем у других. У Hoodia нет
центральных ориентирующих структур и ориентация поллинария кажется
делом случая.
При посещении следующего цветка, опыление
случается, когда ключ поллиния улавливается в отверстие тычиночного
замка и вводится в замок, это происходит в процессе борьбы насекомого,
что бы освободить себя. Зачастую поллинарий ломается у каудикла и
насекомое остаётся с остатком трансляционного аппарата и другим
поллинием. Возможно, что эта сборка опылит следующий цветок, который
посетит это насекомое.
Хотя гибридизация в природе, как известно,
встречается, но она случается нечасто. Имеется несколько факторов
которые, кажется, ответственны за это. Наблюдалось, что опылители
очень специфичны в ограничении себя только лишь одним видом
стапелиевых в тех местах, где можно обнаружить их несколько.
Механические различия являются также лимитирующим фактором в поле.
Ключ черезмерного размера не поместится в более маленький замок. У
Дювалий есть тенденция к наличию большого размера ключа и замка.
Хотя Duvalia и Piaranthus могут быть найдены в одной и той же
местности, они редко гибридизируются. Введение большого ключа
Duvalia предотвращено маленьким замком у Piaranthus и обратное
опыление маловероятно, потому что маленький ключ Piaranthus
проскользнёт сквозь большой замок у Duvalia. В лаборатории,
искустные механические манипуляции делают возможным разместить
поллинии и создать гибрид, который ещё не встречался в
природе. В природе встречаются несколько естественных местообитаний,
где присутствуют механически совместимые виды. Huernia vereckeri и
Huernia longituba ssp. cashelensis растут бок о бок в Зимбабве, и
там же наблюдались многочисленные промежуточные гибриды между ними.
Изготовление большого числа межродовых гибридов искусственным
опылением предполагает, что в этой группе ограничена генетическая
несовместимость. Ограничения полевой гибридизации вызваны, кажется,
прежде всего, факторами географической изоляции, специфичности
опылителей, и механическими барьерами.
Наблюдения поллиний проросших в растворе
глюкозы показали, что местом прорастания пыльцовых трубочек всегда
является ключ. В таком растворе, эта
структура набухает и формирует хорошо различимую пору через которую
пыльцовые трубочки прорастают. (См. фотографию 5). Если
попытаться удалить поллиний в тот же день после введения его в
замок, то обнаружится, что он очень плотно зажат увеличившимся
ключом и развивающимися пыльцовыми трубками. Я полагаю, что это
разбухание ключа, происходящее с поглощением жидкости, удерживает
поллиний на месте и предохраняет его от выталкивания прочь силой
большого количества прорастающих пыльцовых трубочек.
Рассмотрев естественное опыление, давайте теперь
рассмотрим искусственные средства опыления. Хотя, может быть и
возможно развить навыки требуемые для введения поллиния для видов с
большими цветками с помощью ручной лупы, но я считаю, что это
даст очень ограниченную точность сближения.
Вполне вероятно, что визор бинокулярного увеличителя
позволит провести некоторые опыления. Однако, я выяснил, что микроскоп для
препарирования — очень необходимое вспомогательное средство в этой работе. Я использую
Bausch and Lomb Stereozoom инструмент, который позволяет увеличивать
от 10x до 70x крат. Большинство манипуляций было произведено при 20x
кратном увеличении, хотя более маленькие цветки иногда могут
требовать от 40x до 50x. Инструмент крепился на индустриальном
штативе который позволяет ему размещаться над живым высаженным
растением, что бы допустить проведение манипуляций требуемых для
опыления. В начале моих изысканий, я рассматривал вопрос
получения микроманипулятора и косультировался с поставщиками этих
инструментов. Меня уведомили, что они будут иметь слишком
ограниченный диапазон передвижений для моих нужд, но в ходе
обсуждения я узнал, что инструменты используемые в этих
манипуляторах были, как правило, очень тонкие микроиглы сделанные
путём вытягивания из нагретых стеклянных стержней. Я сделал ряд
крючков и игл из вытянутого стекла и нашёл их очень полезными при
манипуляциях поллиниями. К несчастью они были очень
тонкими для этого вида работ, и каждый раз когда я пользовался ими,
они очень быстро ломались. Поиск более подходящих
микро-крючков привёл к открытию очень тонкого #6-0 провода из
нержавеющей стали, который используется, в настоящее время, для
хирургии глаза. Несколько различных крючков было изготовлено с
помощью микроскопа. Они были закреплены в маленьком зажимном патроне
и оставались наиболее важным инструментом для размещения более
мелких поллиниев. Эти крючки были обычно слишком мягкими для удаления
поллиниев, но они были превосходны для ориентации поллиниев и
вставления их в замок. Пинцет для часовщиков #7 является другим
инструментом, который я признал чрезвычайно полезным. Этот пинцет
обычно не достаточно тонок, когда его получаешь, но он может быть
заточен на очень тонком точильном камне при наблюдении под
микроскопом. Очень важно, что бы ровное взаимное расположение
кончиков сохранилось. Пинцет используется для того, что бы сместить
поллинарий и затем удалить его из цветка. Поллинарии помещают на
чистый лист белой бумаги и помечают рисуя круг на бумаге с
наименованием растения находящегося в кругу. Когда
несколько растений используются за одну рабочую сессию, поллинарии
удаляются и помещаются на том же листе бумаги, по мере того, как
каждое растение подносится к микроскопу. Скрещивания делаются, беря
поллинарий с листа бумаги и перенося до нового цветка пинцетом и
помещая его на цветке. Несколько коробочек или небольших круглых
подушек могут использоваться, чтобы стабилизировать рабочую руку.
Все движения под микроскопом должны быть очень плавными движениями
пальцев. Попытка работать без того, чтобы поддерживать руку привела
бы к движениям, которые являются слишком грубыми, чтобы проводить
успешное опыление во всех случаях, кроме больших цветков. Поллиний
затем окончательно располагают с помощью проволочного
крючка, и ключ втягивают в замок. В некоторых случаях необходимо
делать заключительное точное размещение поллиния с помощью пинцета.
Я предпочитаю втягивать поллиний в замок в максимально возможной
степени. Иногда бывают случаи, когда кто-то вставляет маленькие
поллинии в большие замки, и он протягивает поллиний сквозь вершину замка.
С увеличивающимся опытом правильное количество тягового усилия будет
найдено, чтобы предотвратить появления этой ситуации.
Некоторые цветки имеют глубокие трубки венчика,
которые делают доступ к поллиниям затруднённым. Обычно
возможно проделать всю эту работу с более длинным проводом. Когда
это не возможно, венчик может быть отсечен, чтобы открыть доступ к
замку и сделать опыление более простым. Отрезание венчика приводит к
натеканию жидкости из обрезанных кромок. Она может быть поглощена
маленькими кусочками мягкой туалетной бумаги, чтобы предохранить от
влияния влаги сам процесс опыления. Разрезание венчика, кажется, не
оказывает никакого влияния на успех опыления, пока это разрезание не
простирается на те части растения, которые остаются после того, как
цветок обычно отделяется от плодоножки (pedicel).
Встречаются некоторые цветки, которые содержат
большое количествами нектара. И случается, что нектар полностью
покрывает замок, делая искусственное опыление трудным и даже
невозможным. Я выяснил, что его наиболее легко удалить с помощью
маленького заострённого комка туалетной бумаги — прикасаясь им к
нектару с помощью пинцета и наблюдая при этом под микроскопом.
Нектар быстро поглощается и опыление может быть предпринято. Нужно
остерегаться прикасаться к замку этим бумажным комком, поскольку
волокно будет часто застревать в замке и приводить к удалению
поллинария, который может быть необходим для опыления.
В природе, этот лишний нектар, вероятно, удаляется насекомыми,
которые посещают цветок. В опылении Piaranthus, я отметил, что этот
нектар часто полезен, поскольку капиллярное действие жидкости
удерживает поллинарий у поверхности и когда оно снято
ключ скользит в замок очень легко.
Как и с большинством природных процессов, имеются
случаи, когда ошибки в процессе делают невозможным опыление.
Некоторые цветки регулярно демонстрируют поллинии, которые проросли
in situ. Эти поллинии имеют пучок
пыльцевых трубок, выступающих из открытого (разомкнутого) ключа и не
применимы для опыления. Рост этих пыльцевых трубок, кажется,
совершенно случайным и не направленным на замок в любом случае. Эти
цветки не делаются самоопыляющимися от этого in situ прорастания. Я
видел это наиболее часто у Piaranthus, и это может быть связано с
обильным нектаром, упомянутым выше. С другой стороны, цветки
Pseudolithos часто изобилуют нектаром, но я никогда не наблюдал in
situ прорастания у этого рода.
Некоторые клоны регулярно производят цветки с дефектными
поллинариями. Они могут иметь нормальный аппарат транслятора с
присоединенными бесполезными рудиментарными поллиниями. Это повреждение
замечено во всех цветках, произведенных таким клоном. В остальном эти цветки
нормальные и могут быть оплодотворены, но они не могут производить
поллинарии, чтобы оплодотворить другой цветок. В других случаях каудикл
был найден, с такими сильными дефектами, что он не мог ни при каких условиях
транспортировать поллиний. Когда корпускул удален, он отделяется без
присоединенных поллиниев. Как и в другом дефекте, это повреждение
замечено во всех цветках, произведенных этим клоном.
Как только поллиний был вставлен, он
начинает поглощать жидкость из секреций в замке и после 8 или 10
часов, можно видеть очень отчетливое набухание поллиния. Перед
вставкой, поллиний имеет темный, блестящий, глянцевый внешний вид.
После того, как набухание произошло, поллиний становится более светлым,
тусклым и мутным. Можно легко наблюдать прорастание поллиния,
помещая поллиний в нектар находящийся в основании цветка и наблюдая
всё это, на 10 или 12 часов позже, при увеличении.
Имелся некоторый вопрос относительно направления
движения пыльцевых трубок от замка до входа в стручки (follicles). Я
использовал парафиновые срезы опыленных цветов Piaranthus и выяснил,
что эти трубки проникают через губчатую ткань главы рыльца (stigma
head), чтобы достигнуть стручков, которые находятся
непосредственно под ними. В других случаях я препарировал (рассёк)
опыленные цветки из другого рода и увидел пучки пыльцевых трубок,
проникающих через заднюю часть замка, чтобы войти в главу рыльца.
В недавних препарированиях опыленного в природе Asclepias syriaca,
я наблюдал пучки пыльцевых трубочек растущих через место вдоль более
низкого края главы рыльца. Очевидно оба направления роста пыльцевой
трубки встречаются у asclepiads.
В течение двух — четырёх дней после опыления,
венчик отделится от плодоножки (pedicel), и чашелистики закроются над
стручками. Плодоножка, которая не была опылена, может оставаться
присоединённой к цветоножке (peduncle) от нескольких дней до месяца.
Нельзя быть полностью уверенным в удачном опылении до тех пор пока
плодоножка не сохранилась в течение, по крайней мере, месяца. Появление
стручка из фертильной плодоножки происходит через довольно разный период
времени. Самый короткий период, который я наблюдал был одиннадцать
дней. Это было очень удивительно, поскольку, более обычно, что этот процесс
растягивается на многие месяцы или даже годы! Я видел опылённые
образцы Caralluma licatiloba, собранные на Севере Йемена,
образовывавшие плоды с семенами в течение каждого из трёх
последующих лет. В материале, который я искусственно опылил и
вырастил в моей оранжерее, те цветки, которые были
оплодотворены, недавним летом или осенью будет обычно
производить созревшие семена следующей весной. Весенние переопыления
часто дают зрелые семена чуть позже, той же самой весной.
Я должен предостеречь, что сохранение (неопадение)
плодоножки и появление стручков не обязательно гарантируют хороший
результат. Было много случаев, когда производились удалённые
межродовые скрещивания, и когда полностью развившиеся стручки
вырастали, но семена не содержали завязей. Эти семена очень тонкие
на ощупь и имеют вид чешуек которые, конечно же, не будут прорастать.
Поведение плодоножек после успешного опыления различно
и очень интересно. У некоторых видов плодоножка покажет чрезвычайное утолщение.
У других будет происходить большое удлинение, поднимающее стручки намного выше
растения. На тех растениях, которые обычно имеют стелющиеся
плодоножки, плодоножки станут перпендикулярными — поднимая стручки
выше растения. Это поведение плодоножек имеет очевидное значение для
выживания вида, поскольку оно поднимает стручок до уровня, с которого
семена будут более легко рассеяны, когда зрелый стручок раскроется. В
природе семена широко разносятся ветром. Что бы сохранить семена от
потери после раскрывания стручков в оранжерее, я приспособился
использовать трубчатый медицинский напальчник, как наиболее удобный.
Он размещается над созревающими стручками и будет
удерживать семена для того, чтобы их можно было собрать, после их
созревания.
Очень трудно надёжно отгородить цветы в оранжерее
подальше от летающих насекомых. В результате наиболее большие
коллекции стапелиевых демонстрируют частые опыления посредством их.
Семена полученные в результате таких опылений нередко дают интересные
гибриды. Для моих целей такие семена обычно не используются, так как
я имею, более чем достаточно, документированных переопылений для
выращивания. При всех искусственных опылениях, растения защищены -
они содержатся в местах, которые защищены от летающих насекомых или
у них опылены все пять замков так, чтобы опыление насекомыми не
смогло произойти. Все опыления маркируются и записываются для того,
чтобы достоверные данные могли быть накоплены.
Если проблемы культивирования у любителя
выращивающего стапелиевые были полностью решены, прогресс в
получении данных относительно само– и перекрёстной фертильности
(плодовитости), будет намного быстрее. К сожалению, многие
результаты успешных скрещиваний были потеряны из-за болезней растений
в течение трудных зимних месяцев. Это сделало накопление данных в
большом масштабе менее надёжным. В течение периода хорошего роста и
ограниченного ущерба из-за болезней, данные были собраны в ряде
скрещиваний. Эти данные отражены в таблице 5. Можно заметить, что
имеется широкое изменение в степени само-фертильности у различных
родов. Duvalia, Orbea и Stapelianthus
показал самую большую степень само-фертильности. Большинство анализируемых родов показало, по
крайней мере, некоторую само-фертильность. Фертильность у всех родов
гораздо большая, когда проводилось межклональные скрещивание между
членами того же самого вида. Удивительно, но имеются большая степень
фертильности, наблюдаемая при скрещивании между различными видами и
между растениями различных родов. Интересно заметить, что
большинство скрещиваний приводит к гибридизации, дающей растения,
которые являются промежуточными между родителями. В случаях, где
один родитель был полиплоидный — всё потомство имело внешний вид
полиплоидного родителя. Это было отмечено в работе с Orbea woodii
(2n = 44) при скрещиваниях. У большинства стапелиевых, как
сообщалось, число хромосом 2n = 22.
Было отмечено, что у некоторых межродовых гибридов,
потомство имело дефекты в производстве хлорофилла. Сеянцы могут быть
вариегатными и иногда не иметь хлорофилла совсем. В последнем случае
сеянцы умирают вскоре после прорастания. Когда имеется некоторое
количество хлорофилла, растения возможно растут без вариегации. Иногда
получается гибрид, где недостаток хлорофилла постоянен, и светлый
серо-зеленый цвет стебля сохраняется.
Я надеюсь, что эти наблюдения позволят научиться
другим заинтересованным лицам опылять растения из этой необычной
группы. Необходимо некоторое время, чтобы развить требуемые навыки.
Рекомендуется — большая доза терпения.
Табл. 5. Анализ результатов переопылений одного из сезонов.
| Результаты опылений |
| Род |
Самоопылений |
Опылений между клонами |
Опылений между видами |
Опылений межродовых |
| |
Всего сделано |
Процент всхожих |
Всего сделано |
Процент всхожих |
Всего сделано |
Процент всхожих |
Всего сделано |
Процент всхожих |
| Caralluma |
154 |
8% |
25 |
56% |
3 |
67% |
10 |
10% |
| Duvalia |
37 |
22% |
5 |
100% |
5 |
40% |
9 |
33% |
| Echidnopsis |
8 |
0% |
6 |
17% |
--- |
--- |
1 |
0% |
| Frerea |
19 |
0% |
5 |
80% |
--- |
--- |
24 |
13% |
| Huernia |
70 |
6% |
2 |
100% |
18 |
28% |
12 |
50% |
| Orbea |
21 |
29% |
--- |
--- |
34 |
44% |
16 |
17% |
| Piaranthus |
132 |
7% |
18 |
28% |
8 |
100% |
--- |
--- |
| Stapelia |
52 |
4% |
13 |
62% |
14 |
14% |
22 |
19% |
| Stapelianthus |
8 |
38% |
--- |
--- |
4 |
100% |
--- |
--- |
| Tridentia |
16 |
6% |
12 |
58% |
11 |
18% |
6 |
50% |
Благодарности
Я более всего благодарен Darrel
C. H. Plowes, John Lavranos, Inge Pehlemann Brase, Jagdish Sarkaria
и всем другим, которые предоставили мне черенки с документированных
растений, над которыми были выполнены эти исследования. Без этого
материала, многие из этих наблюдений были бы невозможны.
За её терпение, понимание и поддержку в течение тех многих
часов проведенных мною в уединении над микроскопом и фотокамерой, я
хотел бы поблагодарить мою жену, Беатрис.
Ссылки.
- Newton, L. E. 1984. Terminology of structures associated with pollinia
of the Asclepiadaceae. Taxon 33(4):619–621.
- Harold, K. 1985. The worth of pollinarium. Asklepios 35:29.
- Bruyns, P. V. 1985. Pollinaria and Ceropegia arabica and allies. Asklepios 36:65–67.
- Bookman, S. S. 1981. The floral morphology of Asclepias speciosa (Asclepiadaceae)
in relation to pollination and a clarification in terminology for the genus. Amer. J. Bot. 68(5):675–679.
- Schill. R., and Jakel, U. 1978. Beitrag zur Kenninis der Asclepiadaceen–Pollinarien.
Tropische und subtropische Pflanzenwelt 22.
- Schnepf E., Witzig, F., and Schill, R. 1979. Uber Bildung und Feinstruktur des Translators
der Pollinarien von Asclepias curassavica und Gomphocarpus fruticosus.
Tropische und subtropische Pflanzenwelt 25.
- Chaturvedi, S. K. 1987. Pollination and pollen germination in Cynanchium canescens,
Asklepios 40:93–96.
Оригинал статьи расположен по адресу: http://www.cactus-mall.com/stapeliad/pollin.html
Статья опубликована с любезного разрешения автора. Перевод А. Ю. Никитина, Санкт-Петербург, 2006 г.
|